Lejuplādēt prezentāciju
Notiek prezentācijas ielādēšana. Lūdzu uzgaidiet

1
Transkripcija

2
Molekulārās bioloģijas centrālā dogma
Gēni un DNS virza visus dzīves procesus, sniedzot informāciju, kas norāda kā organismam jāattīstās un jāfunkcionē. translācija transkripcija Centrālā dogma apraksta ģenētiskās informācijas plūsmu no DNS uz RNS un proteīnu.

3
Transkripcija Eikariotos DNS nekad nepamet kodolu,
līdz ar to lai DNS nodotu instrukcijas polipeptīdu sintēzei, šai informācijai jābūt nokopētai. Transkripcijas procesā ģenētiskās instrukcijas tiek pārnestas no DNS uz matricas RNS (mRNS).
.")
4
Dzīvnieku šūnās DNS ir sastopama kodolā un mitohondrijos.
Mitohondrijas satur tikai nelielu daļu no kopēja šūnu DNS daudzuma ļoti ierobežotu gēnu skaitu Lielāka daļa šūnas DNS atrodas kodola hromosomās.

5
Gēns ir DNS segments, kas kodē proteīnu vai nekodējošu RNS.
Transkripcijas vienība ir DNS lineāra secība, kas sākās transkripcijas sākuma vietā un beidzās transkripcijas terminācijas (stop) vietā.
 vietā.")
6
A G T C 4 nukleotīdu DNS valoda transkripcija 4 nukleotīdu RNS valoda
proteīnu 20- aminoskābju valoda translācija

7
Tomēr lielāka daļa no šūnas DNS nekad netiek transkribēta.
Kodējošo DNS frakcija genomā ir samērā maza, jo lielāka gēnu sekvences daļa ir nekodējoša; Ievērojama genoma frakcija satur atkārtotas sekvences un funkcionālo gēnu defektīvas kopijas- pseudogēnus un gēnu fragmentus. Šādas sekvences nav funkcionālas un netiek transkribētas mRNS molekulās. Dažādās šūnās transkripcija notiek dažādos DNS fragmentos.

8
Transkripcija ir mRNS sintēze no DNS matricas pavediena

9
Histonu modifikācija un hromatīna remodelēšana
DNS rajoni, kuri netiek transkribēti, atrodas kondensēta hromatīna struktūrā ar ciešu saistību starp histoniem un DNS. Šāda nukleosomu cieša sapakošana neļauj pievienoties proteīniem, kuri mijiedarbojas ar DNS. Histonu modifikācija un hromatīna pārkārtošana var atgriezeniski izmainīt vietējo hromatīna struktūru no kondensētas uz vairāk pieejamu (relaksētu) konformāciju.
 konformāciju.")
10
Eihromatīns un heterohromatīns
Cilvēka genomā izšķir 2 daļas: eihromatīnu (gaišās joslas) un heterohromatīnu (tumšās joslas). Heterohromatīna ir aptuveni 8%, bet eihromatīna ir 92% . Eihromatīns ir samērā pieejams un ir saistīts ar aktīvi transkribētiem gēniem un aktīvi notiekošām histonu modifikācijām. Heterohromatīns ir kondensēts rajons, kurā nenotiek transkripcija, un ir saistīts ar represīvām histonu modifikācijām.
 un. heterohromatīnu (tumšās joslas). Heterohromatīna ir aptuveni 8%, bet eihromatīna ir 92% . Eihromatīns ir samērā pieejams un ir saistīts ar aktīvi transkribētiem gēniem un aktīvi notiekošām histonu modifikācijām. Heterohromatīns ir kondensēts rajons, kurā nenotiek transkripcija, un ir saistīts ar. represīvām histonu modifikācijām.")
11
Histoni Histoni veido nukleosomas tādā veidā , ka to astes ir izbāztas ārā no histonu oktamera. Histonu eksponētas astes satur konservatīvas sekvences. Tie veic svarīgu lomu hromatīna struktūras regulācijā.

12
Histoni Histonu astes sastāda 25- 30% no kopējas histonu masas.
Histonu astes ir bagātas ar Lys (K), Ser (S) un Arg (R), kuri tiek pakļauti modifikācijām. Lys (K): acetilēšana, metilēšana un ubikvitinēšana Arg (R): metilēšana
, Ser (S) un Arg (R), kuri tiek pakļauti modifikācijām. Lys (K): acetilēšana, metilēšana un ubikvitinēšana. Arg (R): metilēšana.")
13
Mehānismi hromatīna struktūras izmainīšanai
Histonu kovalentā modificēšana acetilēšana metilēšana fosforilēšana ubikvitinilēšana sumoilēšana -CH3

14
Histonu kods Modificētās histonu «astes» veido
«histonu kodu», kuru nolasa hromatīnu modificējošie proteīni. Dažādiem «histonu kodiem» var atšķirties bioloģiskās reakcijas iznākums.

15
Histonu acetilēšana Dažādas histonu acetiltransferāzes (histone acetyltransferases, HATs) katalizē acetilgrupu (CH3CO) pievienošanu histonu astēs esošajām lizīna aminogrupām. Lizīna skaits histonu astēs variē no 13 līdz 30. HATs darbojas kā transkripcijas koaktivātori. Kad lādētas Lys sānu ķēdes tiek modificētas tās zaudē savu pozitīvo lādiņu kā rezultātā samazinās saistība starp histoniem un DNS pie tām saistās hromatīna remodelējošie kompleksi. RNS Pol un transkripcijas faktoriem ir vieglāk piekļūt promotera rajonam.
 katalizē acetilgrupu (CH3CO) pievienošanu histonu astēs esošajām lizīna aminogrupām. Lizīna skaits histonu astēs variē no 13 līdz 30. HATs darbojas kā transkripcijas koaktivātori. Kad lādētas Lys sānu ķēdes tiek modificētas. tās zaudē savu pozitīvo lādiņu kā rezultātā samazinās saistība starp histoniem un DNS. pie tām saistās hromatīna remodelējošie kompleksi. RNS Pol un transkripcijas faktoriem ir vieglāk piekļūt promotera rajonam.")
16
Histonu deacetilēšana
Histonu deacetilāzes (HDACs) noņem acetil grupas no Lys atlikumiem un veicina transkripcijas nomākšanu. HDACs tiek piesaistītas kā daļa no korepresora kompleksa. Pastiprina mijiedarbību starp DNS un histoniem.
 noņem acetil grupas no Lys atlikumiem un veicina transkripcijas nomākšanu. HDACs tiek piesaistītas kā daļa no korepresora kompleksa. Pastiprina mijiedarbību starp DNS un histoniem.")
17
Histonu metilēšana Histonu metiltransferāzes (HMTs) ir enzīmi, kas katalizē metīlgrupu pārnesi uz histonu proteīna Arg (R) un Lys (K) atlikumiem. Histonu metilēšana var izraisīt dažādu faktoru piesaistīšanos un transkripciju gan aktivēt, gan nomākt (represēt). Histonu Arg metilēšana ir iesaistīta transkripcijas aktivācijā. Histonu Lys metilēšana var represēt transkripciju. Metilēšana var būt mono-, di-, vai tri-metilēta. Monometilēšana ir saistīta ar gēnu aktivāciju, savukārt trimetilēšana ir saistīta ar gēna represiju.
. Histonu Arg metilēšana ir iesaistīta transkripcijas aktivācijā. Histonu Lys metilēšana var represēt transkripciju. Metilēšana var būt mono-, di-, vai tri-metilēta. Monometilēšana ir saistīta ar. gēnu aktivāciju, savukārt. trimetilēšana ir saistīta ar gēna represiju.")
18
Hromatīna remodelējošie kompleksi
Sastāv no daudzām (>10) subvienībām. Papildus ATPāzes domēniem, tie satur arī domēnus, kuri mijiedarbojas ar modificētiem histoniem. Tas nodrošina saskaņotu darbību starp histonu modificēšanu un hromatīna remodelēšanu. satur bromodomēnus, kas mijiedarbojas ar histonu acetilētiem Lys atlikumiem satur hromodomēnus, kas mijiedarbojas ar histonu metilētiem Lys atlikumiem
 subvienībām. Papildus ATPāzes domēniem, tie satur arī domēnus, kuri mijiedarbojas ar modificētiem histoniem. Tas nodrošina saskaņotu darbību starp histonu modificēšanu un hromatīna remodelēšanu. satur bromodomēnus, kas mijiedarbojas ar histonu acetilētiem. Lys atlikumiem. satur hromodomēnus, kas mijiedarbojas ar. histonu metilētiem Lys atlikumiem.")
19
Hromatīna remodelēšana
Hromatīna remodelēšanas (proteīnu) komplekss īslaicīgi maina nukleosomu struktūru un izvietošanu, izmantojot ATP hidrolīzes reakcijas enerģiju. Hromatīna remodelēšanas faktori ir multiproteīnu kompleksi, kas satur dažas subvienības ar helikāzes aktivitāti. Remodelēšana padara pieejamas DNS vai histonu piesaistes vietas transkripcijas regulātoriem vai histonu modificējošiem proteīniem. Kompleksa aktivitāte var izraisīt arī transkripcijas represiju, jo dekondensētais hromatīns ir pieejamāks arī histonu deacetilāzēm.
 komplekss īslaicīgi maina nukleosomu struktūru un izvietošanu, izmantojot ATP hidrolīzes reakcijas enerģiju. Hromatīna remodelēšanas faktori ir multiproteīnu kompleksi, kas satur dažas subvienības ar helikāzes aktivitāti. Remodelēšana padara pieejamas DNS vai histonu piesaistes vietas transkripcijas regulātoriem vai histonu modificējošiem proteīniem. Kompleksa aktivitāte var izraisīt arī transkripcijas represiju, jo dekondensētais hromatīns ir pieejamāks arī histonu deacetilāzēm.")
20
Hromatīna remodelēšana
Remodelēšanas rezultātā nukleosoma var mainīt savu pozīciju, slīdot gar DNS. Nukleosomas slīdēšanas rezultātā, DNS sekvence, kura sākumā bija aptīta apkārt histonu oktamēriem, kļūst pieejama transkripcijas mašinērijai.

21
Kas ir nepieciešams transkripcijai?
dsDNS matrica promoters = transkripcijas uzsākšanas vieta ribonukleotīdi RNS polimerāze transkripcijas faktori

22
Transkripcijas posmi Eikariotu transkripcija notiek trijos secīgos posmos: Iniciācija ir transkripcijas sākums. Enzīms RNS polimerāze (RNS Pol) piesaistās pie gēna rajona, ko sauc par promoteru. DNS saņem signālu atvīt pavedienus. 2. Elongācija ir nukleotīdu pievienošana mRNS pavedienam. RNS Pol nolasa DNS pavedienu un sintezē mRNS, izmantojot komplementārus bāzu pārus. 3. Terminācija ir transkripcijas apturēšana. RNS Pol sasniedz gēna stop/terminācijas sekvenci un atdalās no DNS.
 piesaistās pie gēna rajona, ko. sauc par promoteru. DNS saņem signālu atvīt pavedienus. 2. Elongācija ir nukleotīdu pievienošana mRNS pavedienam. RNS Pol nolasa DNS pavedienu un sintezē mRNS, izmantojot. komplementārus bāzu pārus. 3. Terminācija ir transkripcijas apturēšana. RNS Pol sasniedz gēna stop/terminācijas sekvenci un atdalās no DNS.")
23
ribosomāliem RNS (rRNS):
RNS polimerāzes Eikariotu šūnai ir 3 dažādas kodola RNS polimerāzes. Tās transkribē dažādu klašu gēnus. RNS polimerāze I RNS polimerāze II RNS polimerāze III pre-rRNS 45S, kas ir prekursors 3 ribosomāliem RNS (rRNS): 28S* 18S 5.8S Proteīnus kodējošie gēni daži snRNAs mikro RNS Transporta RNS (tRNS) gēni 5S rRNS (mazākā ribosomālā RNS) Mazās kodola RNS (snRNA) iesaistītas splaisingā Mazās citoplazmas (Small cytoplasmic (scRNS) iesaistītas proteīna transportā *rRNA S ir Svedberga vienība, ko izmanto molekulāro struktūru izmēru raksturošanai ar ultracentrifugācijas palīdzību.
: 28S* 18S. 5.8S. Proteīnus. kodējošie gēni. daži snRNAs. mikro RNS. Transporta RNS (tRNS) gēni. 5S rRNS (mazākā ribosomālā. RNS) Mazās kodola RNS (snRNA) iesaistītas splaisingā. Mazās citoplazmas (Small cytoplasmic (scRNS) iesaistītas proteīna. transportā. *rRNA S ir Svedberga vienība, ko izmanto molekulāro struktūru. izmēru raksturošanai ar ultracentrifugācijas palīdzību.")
24
ribosomāliem RNS (rRNS):
RNS polimerāzes RNS polimerāze I RNS polimerāze II RNS polimerāze III Transporta RNS (tRNS) gēni 5S rRNS (mazākā ribosomālā RNS) Mazās kodola RNS (snRNA) iesaistītas splaisingā Small cytoplasmic (scRNS) iesaistītas proteīna transportā pre-rRNS 45S, kas ir prekursors 3 ribosomāliem RNS (rRNS): 28S* 18S 5.8S Proteīnus kodējošie gēni daži snRNAs mikro RNS RNS Pol I transkribē 1 gēnu ≈ 200 kopijās. RNS Pol II transkribē ≈ gēnus. RNS Pol III transkribē gēnus variablā kopiju skaitā.
 gēni. 5S rRNS (mazākā ribosomālā. RNS) Mazās kodola RNS (snRNA) iesaistītas splaisingā. Small cytoplasmic (scRNS) iesaistītas proteīna. transportā. pre-rRNS 45S, kas. ir prekursors 3. ribosomāliem RNS (rRNS): 28S* 18S. 5.8S. Proteīnus. kodējošie gēni. daži snRNAs. mikro RNS. RNS Pol I transkribē 1 gēnu ≈ 200 kopijās. RNS Pol II transkribē ≈ gēnus. RNS Pol III transkribē gēnus variablā kopiju skaitā.")
25
RNS polimerāzes Visas trīs kodola RNS polimerāzes ir kompleksie enzīmi. Katra RNS Pol sastāv no 8 līdz 17 dažādām subvienībām. Neskatoties uz to, ka tās atpazīst dažādus promoterus, transkribē dažādu klašu gēnus, visām polimerāzēm ir kopīgas iezīmes.

26
RNS polimerāze I un III Rauga RNS Pol I struktūra tika atrisināta gadā. RNS Pol I ir 590 kDa enzīms, kurš sastāv no 14 subvienībām. No tām 12 subvienībām ir identiskas vai līdzīgas daļas ar RNS Pol II un III subvienībām. RNS Pol III sastāv no 17 subvienībām.

27
RNA polymerase II RNS Pol II ir 550 kDa enzīms, kuru veido 12 subvienības. 12 subvienības kopā veido krabja spīlei līdzīgo formu. Žokļi (jaws) satver DNS matricu un skavas (clamp) notur matricu polimerāzes katalītiskā centrā lai veicinātu mRNS sintēzi. Katalītiskais centrs satur Mg2+ jonus.
 satver DNS matricu un skavas (clamp) notur matricu polimerāzes katalītiskā centrā lai veicinātu. mRNS sintēzi. Katalītiskais centrs satur Mg2+ jonus.")
28
RNS polimerāzei II veidojas vairāki
kanāli, kas nodrošina DNS, RNS un nukleotīdu virzību uz katalītisko centru un ārā no tā.

29
Transkripcijas faktori
Eikariotu RNS Pol pašas pa sevi nespēj iesākt transkripcijas procesu. RNS Pol II nespēj atpazīt promotera sekvences un atvīt DNS dubultspirāli. Līdz ar to transkripcijas iniciācijai ir nepieciešami specifiskie proteīni- transkripcijas faktori. Pastāv divu veida transkripcijas faktori: Vispārēji transkripcijas faktori ir iesaistīti visās transkripcijās, kas sākās RNS Pol promoteros. Tie ir sastāvdaļa no galvenās transkripcijas mašinērijas. Papildus transkripcijas faktori piesaistās pie DNS sekvencēm, kas kontrolē individuālo gēnu ekspresiju un atbild par gēnu ekspresijas regulāciju.

30
Regulatorie elementi un transkripcijas faktori
Īso sekvenču elementu kombinācija katra gēna tuvumā darbojas kā transkripciju faktoru atpazīšanas signāli (regulatorie elementi). Lielākā daļa no īso sekvenču elementiem atrodas pirms gēna kodējošas sekvences, kur tie kopā veido promoteru. Transkripciju faktoru atpazīšanas signāli (promoteru elementi) ir cis-iedarbības, jo tie darbojās tajā pašā DNS vietā, kur paši atrodas. Transkripcijas faktori ir trans- iedarbības, jo tos sintezē gēni, kas atrodas tālu no promotera vietas un tiem ir jāmigrē uz savu darbības vietu.
. Lielākā daļa no īso sekvenču elementiem atrodas pirms gēna kodējošas sekvences, kur tie kopā veido promoteru. Transkripciju faktoru atpazīšanas signāli (promoteru elementi) ir cis-iedarbības, jo tie darbojās tajā pašā DNS vietā, kur paši atrodas. Transkripcijas faktori ir trans- iedarbības, jo tos sintezē gēni, kas atrodas tālu no promotera vietas un tiem ir jāmigrē uz savu darbības vietu.")
31
Vispārējie transkripcijas faktori (GTFi, General transcription factors)
Nepieciešami RNS polimerāzes piesaistei un transkripcijas uzsākšanai no lielākās daļas promoteru Prokariotos ir tikai viens GTF – sigma faktors Eikariotos ir vismaz 6 GTFi – TFIIA, TFIIB, TFIID, TFIIE, TFIIF un TFIIH

32
Promoteri Promoteri atrodas augšup gēnam.
Promoteru garums var būt no 100 līdz 1000 bāzu pāriem. Visbiežākie cis-iedarbības promoteru elementi: TATA bokss , biežāk TATAAA. Parasti tas atrodas bāzu pārus pirms transkripcijas sākuma vietas (-25). Tikai 10-15% zīdītāju gēni satur TATA boksu, pārējiem ir citi promoteru elementi. GC bokss , biežāk GGGCGG, ir sastopams daudzos gēnos, parasti tajos kuriem nav TATA bokss. CAAT bokss, bieži atrodas pozīcijā -80.
. Tikai 10-15% zīdītāju gēni satur TATA boksu, pārējiem ir citi promoteru elementi. GC bokss , biežāk GGGCGG, ir sastopams daudzos gēnos, parasti tajos kuriem nav TATA bokss. CAAT bokss, bieži atrodas pozīcijā -80.")
33
Eikariotiskie promoteri TATA bokss
Vislabāk izpētītā promoteru sekvence Atrodas daudzu, bet ne visu gēnu 5’ UTR sastāvā nukleotīdus pirms transkripcijas sākuma vietas Piesaista t.s. TATA boksa piesaistīšanās proteīnu (TBP), kas ir transkripcijas faktora TFIID sastāvdaļa
, kas ir transkripcijas faktora TFIID sastāvdaļa.")
34
Citi eikariotisko promoteru elementi
-100 līdz -1 -37 līdz -32 -31 līdz -26 -2 līdz +4 +28 līdz +32 CpG BRE TATA Inr DPE GGG CCA A T T A G A C A G AC TT CGCG TATA AA PyPyAN PyPy G CpG salas – CG bagātas sekvences, piedalās transkripcijas regulācijā, C nukleotīds var būt metilēts BRE (TFII B recognition element) – tieši piesaista transkripcijas faktoru IIB Iniciatora elements (Inr) darbojas līdzīgi TATA boksam un arī piesaista TBP DPE elements (Downstream promoter element) piesaista citas TFIID subvienības- TAF9 un TAF5 Eikariotisko promoteru sastāvā var būt viens vai vairāki no minētajiem elementiem
 – tieši piesaista transkripcijas faktoru IIB. Iniciatora elements (Inr) darbojas līdzīgi TATA boksam un arī piesaista TBP. DPE elements (Downstream promoter element) piesaista citas TFIID subvienības- TAF9 un TAF5. Eikariotisko promoteru sastāvā var būt viens vai vairāki no minētajiem elementiem.")
35
Enhānseri Enhānseri ir 50 - 150 bp garuma DNS fragmenti.
Tie ir cis-iedarbības sekvences, kas paaugstina daudzu promoteru darbību. Enhānseri veic stimulējošo darbību uz promotoriem paši būdami vairāku tūkstošu bāzu pāru attālumā no promotera. Var atrasties un darboties jebkurā vietā- pirms gēna, pēc gēna vai intronā. Tie saistās ar gēna regulatoriem elementiem, līdz ar to DNS starp promoteru un enhanseri veido cilpu. Tas ļauj pie enhānseriem piesaistītiem proteīniem mijiedarboties ar promotera transkripcijas faktoriem.

36
Transkripcijas iniciācija eikariotos - pārskats
Enhancer

37
Iniciācijas posms Polimerāzes – promoteru kompleksa struktūrā notiek
strukturālas izmaiņas lai iesāktu transkripciju. Transkripcijas vietā aptuveni DNS 14 bāzu pāri atvijas un RNS Pol II ar Vispārējiem transkripcijas faktoriem veido transkripcijas “burbuli”. mRNS sintēze notiek 5’- 3’ virzienā (pieaug 3’- galā).
.")
38
Preiniciācijas komplekss
Vispārēji transkripcijas faktori (GTF) savienojās ar RNS Pol veidojot preiniciācijas kompleksu. Preiniciācijas komplekss Spēj iniciēt transkripcijas procesu Palīdz novietot RNS Pol II virs gēna transkripcijas starta vietas Denaturē DNS (atdala DNS pavedienus)
 savienojās ar RNS Pol veidojot. preiniciācijas kompleksu. Preiniciācijas komplekss. Spēj iniciēt transkripcijas procesu. Palīdz novietot RNS Pol II virs gēna transkripcijas starta vietas. Denaturē DNS (atdala DNS pavedienus)")
39
Preiniciācijas komplekss
Preiniciācijas komplekss satur : RNS Pol II 6 vispārējos transkripcijas faktorus (TF) TFIIB TFIIA Koaktivātoru komplekss TFIID TFIIE RNS Pol un vispārēji TF piesaistās pie iniciācijas kompleksa noteiktā secībā . TFIIH TFIIF
 TFIIB. TFIIA. Koaktivātoru komplekss. TFIID. TFIIE. RNS Pol un vispārēji TF piesaistās pie. iniciācijas kompleksa noteiktā secībā . TFIIH. TFIIF.")
40
Transkripcijas iniciācija
TATA piesaistes proteīns (TBP) ir daļa no TFIID kompleksa. Tam ir jāpiesaistās pie promotera vēl pirms citiem vispārējiem TF un RNS Pol II. Lielākai daļai gēnu nav TATA boksa, līdz ar to tiek izmantots iniciātora elements (Inr) vai kāds no citiem promoteriem. TBP piesaistās nespecifiski, pievienojot atlikušās TFIID subvienības pie promotera.
 ir daļa no TFIID kompleksa. Tam ir jāpiesaistās pie promotera vēl pirms citiem vispārējiem TF un RNS Pol II. Lielākai daļai gēnu nav TATA boksa, līdz ar to. tiek izmantots iniciātora elements (Inr) vai kāds no citiem promoteriem. TBP piesaistās nespecifiski, pievienojot atlikušās TFIID. subvienības pie promotera.")
41
RNS Pol un GTF veido preiniciācijas kompleksu īpašā secībā
TFIIA mijiedarbojas ar TFIID faktora TBP subvienību, veidojot DA kompleksu. Tas veicina preiniciācijas kompleksa veidošanos. Nākamais piesaistās TFIIB veidojot DAB kompleksu. TFIIB novieto DNS pareizajā pozīcijā, lai tas varētu ieiet RNS Pol II aktīvajā saitā. DAB komplekss piesaista RNS Pol II un TFIIF. RNS Pol II ar TFIIF palīdzību piesaistās rajonā no -34 bp līdz +17bp, veidojot DABPol IIF kompleksu. TFIIF paātrina polimerizācijas procesu.

42
RNS Pol un GTF veido preiniciācijas kompleksu īpašā secībā
Pēdējie pievienojās TFIIE un TFIIH, veidojot pilnu preiniciācijas kompleksu- DABPolIIFEH. TFIIE palīdz atvērt/aizvērt RNS Pol II krabja spīlei līdzīgo struktūru, kas ļauj polimerāzei pa DNS pavedienu virzīties uz priekšu.

43
RNS Pol un GTF veido preiniciācijas kompleksu īpašā secībā
Divas subvienības TFIIH ir helikāzes, kuras atvij DNS pavedienu transkripcijas iniciācijas vietā, veidojot transkripcijas burbuli. Transkripcijas burbulis ir molekulārā struktūra, kas veidojas DNS transkripcijas laikā, kad neliels DNS spirāles fragments ir atvīts. Viena no TFIIH subvienībām ir proteīna kināze, kas fosforilē RNS Pol II C-terminālajā domēnā (CTD) esošās aminoskābes- serīnus. Tas stimulē RNS Pol II uzsākt mRNS sintēzi.
 esošās aminoskābes- serīnus. Tas stimulē RNS Pol II uzsākt mRNS sintēzi.")
44
Koaktivātoru komplekss
Mediators ir multiproteīnu komplekss, kas darbojas kā transkripcijas koaktivātors. Mediators veido tiltu starp aktivātoriem (Act), kas ir piesaistīti pie DNS regulatoriem elementiem (RE), un galveno transkripcijas mašinēriju (Pol II un GTF). Mediatora kompleksu veido 31 subvienība un to var iedalīt 4 daļās – galva, vidus, aste un disociējoša kināze. Mediators piesaistās pie RNS Pol II C-terminālā domēna (CTD) un veido tiltu starp šo fermentu un transkripcijas faktoru. Koaktivātori ir nepieciešami daudzu regulatoro signālu sasaistei. Pēdējo gadu pētījumi sasaistīja Mediatorā sastopamās mutācijas ar dažādām slimībām, ieskaitot Alchemera slimību, vēzi un iedzimtos izkropļojumus.
, kas ir piesaistīti pie DNS regulatoriem elementiem (RE), un. galveno transkripcijas mašinēriju (Pol II un GTF). Mediatora kompleksu veido 31 subvienība un to var iedalīt 4 daļās – galva, vidus, aste un disociējoša kināze. Mediators piesaistās pie RNS Pol II C-terminālā domēna (CTD) un veido tiltu starp šo fermentu un transkripcijas faktoru. Koaktivātori ir nepieciešami daudzu regulatoro signālu sasaistei. Pēdējo gadu pētījumi sasaistīja Mediatorā sastopamās mutācijas ar dažādām slimībām, ieskaitot Alchemera slimību, vēzi un iedzimtos izkropļojumus.")
45
SAGA SAGA (Spt-Ada-Gcn5 acetiltransferāze) ir multi-proteīnu hromatīnu modificējošais Komplekss, kas un darbojas kā transkripcijas koaktivātors. SAGA atver hromatīnu transkripcijas faktoru un preiniciācijas kompleksa piesaistei.

46
Elongācijas posms Transkripcijas process pāriet no iniciācijas posma elongācijas posmā pēc tam, kad RNS Pol ir uzsintezējusi īsu (10 nukleotīdu) mRNS fragmentu. Šajā transkripcijas posmā RNS Pol II notiek konformacionālas izmaiņas kuru rezultātā Pol stiprāk satver DNS matricu.
 mRNS fragmentu. Šajā transkripcijas posmā RNS Pol II notiek konformacionālas izmaiņas. kuru rezultātā Pol stiprāk satver DNS matricu.")
47
Elongācijas posms RNS Pol II CTD (C-terminālā domēna) fosforilācija ļauj atdalīt RNS Pol II no vispārējiem TF un uzsākt transkripcijas elongācijas fāzi. Elongācijas laikā, TFIIF (piemīt helikāzes aktivitāte) paliek savienota ar RNS Pol II, bet visi pārējie TF disociē no preiniciācijas kompleksa. RNS Pol izmanto NTPs RNS transkripta sintēzei. RNS Pol (līdzīgi DNS Pol) var pievienot nukleotīdus tikai pavediena 3‘ galā. RNS ir jāsintezē 5' - 3' virzienā. Tikai viens no atvītiem DNS pavedieniem kļūs par matricu mRNS sintēzei. Transkribēta DNS atjauno dubultspirāli.
 fosforilācija ļauj atdalīt RNS Pol II no vispārējiem TF un uzsākt transkripcijas elongācijas fāzi. Elongācijas laikā, TFIIF (piemīt helikāzes aktivitāte) paliek savienota ar RNS Pol II, bet visi pārējie TF disociē no preiniciācijas kompleksa. RNS Pol izmanto NTPs RNS transkripta sintēzei. RNS Pol (līdzīgi DNS Pol) var pievienot nukleotīdus tikai pavediena 3‘ galā. RNS ir jāsintezē virzienā. Tikai viens no atvītiem DNS pavedieniem kļūs par matricu mRNS sintēzei. Transkribēta DNS atjauno dubultspirāli.")
48
Terminācijas posms Eikariotu proteīnu gēni satur poli-A signālu lejup no pēdēja eksona. Transkripcija bieži terminējās kb lejup no poli-A signāla, bet precīzs mehānisms vēl nav zināms. Kad transkripcija terminējās, mRNS Pol un mRNA transkripts atbrīvojās no DNS. DNS dubultspirāle atjaunojās.

49
Pēc transkripcijas... Prokariotos mRNS ir gatava translācijai uzreiz pēc transkripcijas vai pat vienlaicīgi ar to. Eikariotos vispirms ir jāveic mRNS procesēšana un transports uz citoplazmu.

50
Pēctranskripcijas modifikācijas mRNS procesēšana
Transkripcijas procesā izveidotais RNS ir proteīnu kodējošo gēnu primārais transkripts. Tas ir prekursoru mRNS (pre-mRNS). Pre- mRNS ir pakļauts daudzām izmaiņām, tā saucamām pēctranskripcijas modifikācijām (RNS procesēšanai). RNS procesēšana ir nepieciešama, lai izveidotu funkcionālas mRNS molekulas. Šāda mRNS molekula tiek transportēta uz citoplazmu, kur tiek tālāk translēta par proteīnu.
. Pre- mRNS ir pakļauts daudzām izmaiņām, tā saucamām pēctranskripcijas. modifikācijām (RNS procesēšanai). RNS procesēšana ir nepieciešama, lai izveidotu funkcionālas mRNS. molekulas. Šāda mRNS molekula tiek transportēta uz citoplazmu, kur tiek tālāk translēta. par proteīnu.")
51
pre-mRNS procesēšana 1. Kepings (cappings) 2. Poliadenilēšana
3. Splaisings

52
RNS kepings Kepings notiek uzreiz pēc mRNS pavediena
5’ gala nokāpšanas no RNS Pol II.

53
RNS kepings Uzreiz pēc transkripcijas metilētais nukleozīds, 7-metilguanozīns (m7G), tiek pievienots RNS transkripta 5’ galā esošajam nukleotīdam ar 5’- 5’ fosfodiestera saitu. 5’-5’ triphosphate bridge RNS kepings nobloķē RNS transkripta 5’ galu.

54
RNS kepings Kepinga funkcijas: pasargā mRNS transkriptu no 5’- 3’
eksonukleāzes uzbrukuma. mRNA bez kepa tiek ātri degradētas atvieglo mRNS transportu no kodola uz citoplazmu nepieciešams RNS splaisingam spēlē būtisku lomu ribosomālās 40S subvienības piesaistē pie mRNA (ir nepieciešams mRNS translācijai).
.")
55
RNS poliadenilēšana Praktiski visām mRNS molekulām 3’- galā ir vairāki adenilāta atlikumi– poli(A)-aste. Šie daudzi A netiek kodēti gēnos. Pre-mRNS 3’- galā tiek sašķelts noteiktā vietā un poli(A) polimerāze pievieno adenilāta atlikumus.
 polimerāze pievieno adenilāta atlikumus.")
56
RNS poliadenilēšana Šķelšanas un poliadenilēšanas reakcija notiek, ja
galvenā poliadenilēšanas signāla (PAS) sekvence - 5'- AAUAAA-3' atrodas tuvu pre-mRNS molekulas 3‘- galam. Šķelšana notiek vietā, kas atrodas nukleotīdus lejup no AAUAAA elementa, parasti tā ir 5'-CA-3‘ sekvence. G/U-bagāta sekvence (Downstream Signal Element) savukārt atrodas lejup no pre-mRNS šķelšanas vietas.
 sekvence AAUAAA-3 atrodas tuvu pre-mRNS molekulas 3‘- galam. Šķelšana notiek vietā, kas atrodas nukleotīdus lejup no AAUAAA elementa, parasti tā ir 5 -CA-3‘ sekvence. G/U-bagāta sekvence (Downstream Signal Element) savukārt atrodas lejup no. pre-mRNS šķelšanas vietas.")
57
RNS poliadenilēšana Divi multisubvienību proteīni:
Šķelšanas un poliadenilēšanas specifiskais faktors (Cleavage and Polyadenylation Specificity Factor (CPSF)) un Šķelšanas stimulācijas factors (Cleavage Stimulation Factor (CStF)) tiek pārnesti no RNS Pol II uz mRNS molekulu un saistās ar sekvences elementiem. Šis komplekss šķeļ mRNS šķelšanas vietā 5'-CA-3‘, kas atrodas starp PAS un G/U-bagāto sekvenci.
) un. Šķelšanas stimulācijas factors. (Cleavage Stimulation Factor (CStF)) tiek pārnesti no RNS Pol II uz mRNS molekulu un. saistās ar sekvences elementiem. Šis komplekss šķeļ mRNS šķelšanas vietā 5 -CA-3‘, kas atrodas starp PAS un G/U-bagāto sekvenci.")
58
RNS poliadenilēšana Pēc tam poliadenilāta polimerāze (PAP) jaunajam mRNA molekulas 3’ galam pievieno aptuveni 200 adenīna vienības, izmantojot adenozīna trifosfātu (ATP) kā prekursoru. Tomēr, poliadenilāta polimerāze vāji turās pre-mRNS galā kamēr nav pievienotas vismaz 20 adenīna (A) vienības. Tad poli(A)-piesaistes proteīns (PAB) piesaistās pie īsās poli(A) astes un palīdz poliadenilāta polimerāzei stingrāk piesaistīties. Tas arī veicina ātru A vienību pievienošanu. PAB proteīns arī pasargā mRNS 3’ galu no ribonukleāzes darbības (šķelšanas).
 jaunajam mRNA molekulas 3’ galam pievieno aptuveni 200 adenīna vienības, izmantojot adenozīna trifosfātu (ATP) kā prekursoru. Tomēr, poliadenilāta polimerāze vāji turās pre-mRNS galā kamēr nav pievienotas vismaz 20 adenīna (A) vienības. Tad poli(A)-piesaistes proteīns (PAB) piesaistās pie īsās poli(A) astes un palīdz poliadenilāta polimerāzei stingrāk piesaistīties. Tas arī veicina ātru A vienību pievienošanu. PAB proteīns arī pasargā mRNS 3’ galu no ribonukleāzes darbības (šķelšanas).")
59
RNS poliadenilēšana RNS poliadenilēšana ir nepieciešama:
transkripcijas terminēšanai mRNS molekulas transportam uz citoplazmu lai pasargātu mRNS molekulu no tās degradācijas citoplazmā (mRNA degradācija ir saistīta ar poli(A) astes saīsināšanu) translācijas procesam, paaugstinot ribosomālās mašinērijas piesaisti pie mRNA molekulas.
 astes saīsināšanu) translācijas procesam, paaugstinot ribosomālās mašinērijas. piesaisti pie mRNA molekulas.")
60
Eksoni, introni un gēnu gali
Introni – nekodējošās sekvences gēnu vidus daļā Eksoni – gēnu kodējošās sekvences Introni bieži tiek uzlūkoti kā “nederīga” DNS, bet tie var saturēt regulatoras sekvences vai funkcionālas nekodējošās RNS Parasti introni ir daudz garāki par eksoniem Introni ir sastopami eikariotos, prokariotos tie ir daudz retāki Gēnu 5’un 3’ gali satur t.s. netranslētos reģionus (UTR), kuri satur regulējošas sekvences, piem. promoteri
, kuri satur regulējošas sekvences, piem. promoteri.")
61
pre-mRNS splaisings Transkripcijas procesā sākumā izveidojas pre-mRNS sekvence, kas ir komplementāra pilna garuma gēnam. Splaisinga procesā no transkribētas pre-mRNS sekvences tiek izgriezti introni un savā starpā savienoti eksoni. Tā rezultātā izveidojas mRNS molekula.

62
pre-mRNS splaisinga regulatorie elementi
Pre-mRNS satur “splaisinga kodu”, kuru veido konsensus sekvences, kas nosaka splaisinga savienošanas vietas samērā īsi cis-darbības elementi eksonos vai intronos.

63
pre-mRNS splaisinga regulatorie elementi
mRNA spalisingam ir nepieciešamas īpašas nukleotīdu sakvences eksona/introna savienojuma vietā. Vairumā gadījumu, introns sākās no GU sekvences (5' splaisa vieta) un beidzās ar sekvenci AG (3' splaisa vieta) - GT-AG likums). Minētas sekvences ir ļoti svarīgas splaisingam, bet ir nepietiekamas. Trešā svarīgā vieta splaisingā ir konservatīvais A nukleotīds. Tas parasti atrodas ļoti tuvu introna beigām, aptuveni 20–50 bāzes pirms AG (3' splaisa vietas) nukleotīdiem.
 un beidzās ar sekvenci AG (3 splaisa vieta) - GT-AG likums). Minētas sekvences ir ļoti svarīgas splaisingam, bet ir nepietiekamas. Trešā svarīgā vieta splaisingā ir konservatīvais A nukleotīds. Tas parasti atrodas ļoti tuvu introna beigām, aptuveni 20–50 bāzes pirms AG (3 splaisa vietas) nukleotīdiem.")
64
RNS splaisings Splaisingu katalizē RNS/proteīnu komplekss-
splaiseosoma. Splaiseosoma ir komplekss, kas sastāv no piecu veida mazām nukleolārām RNS ( nt small nuclear RNA (snRNA)) un vairāk nekā 50 proteīniem. Katra no snRNA molekulām ir piesaistīta pie proteīniem un veido mazos nukleāros ribonukleoproteīnus- snurps (snRNPs, small nuclear ribonucleoproteins). snRNP -U1, U2, U4, U5 and U6. snRNPs piedalās spalisinga procesā atpazīstot mRNS sekvences vai mainot splaiseosomas konformāciju. Splaiseosomas katalizē reakciju, kuras rezultātā tiek izgriezti introni un «salīmēti" kopā kodējošie eksoni.
) un vairāk nekā 50 proteīniem. Katra no snRNA molekulām ir piesaistīta pie. proteīniem un veido mazos nukleāros ribonukleoproteīnus- snurps (snRNPs, small nuclear ribonucleoproteins). snRNP -U1, U2, U4, U5 and U6. snRNPs piedalās spalisinga procesā atpazīstot mRNS sekvences vai mainot splaiseosomas. konformāciju. Splaiseosomas katalizē reakciju, kuras rezultātā tiek izgriezti introni un «salīmēti kopā kodējošie eksoni.")
65
pre-mRNS splaisings snRNPs atpazīst un piesaistās pie
introna abiem 5’ un 3’ galiem kā rezultātā introna gali satuvinās, veidojot «laso" struktūru. Šķelšana 3’ savienojuma galā, noved pie introna izgriešanas un eksonu savienošanas.

66
pre-mRNS splaisinga regulatorie elementi
Papildus, splaicingam nepieciešamā informācija atrodas samērā īsās (~6 nukleotīdu) sekvencēs, kas ir eksonos un intronos un var gan pastiprināt, gan nomākt splaisingu. Eksonu splaicinga enhānseri (ESEs) un eksonu splaicinga klusinātāji (ESSs) . Intronu splaicinga enhānseri(ISEs) and intronu splaicinga klusinātāji (ISSs). Atšķirībā no konsensus spalisinga vietām, šiem regulatoriem elementiem nav noteiktas sekvences un pozīcijas.
 sekvencēs, kas ir eksonos un intronos un var gan pastiprināt, gan nomākt splaisingu. Eksonu splaicinga enhānseri (ESEs) un eksonu splaicinga klusinātāji (ESSs) . Intronu splaicinga enhānseri(ISEs) and intronu splaicinga klusinātāji (ISSs). Atšķirībā no konsensus spalisinga vietām, šiem regulatoriem elementiem nav noteiktas sekvences un pozīcijas.")
67
pre-mRNS splaisinga regulatorie elementi
Divu veida trans- darbības splaisinga faktori var mijiedarboties ar enhānseriem un klusinātajiem: serīna arginīna (SR) saimes proteīni un heterogēnās ribonucleoproteīna daļiņas (hnRNPs). Cilvēka nRNP proteīni satur vienu vai vairākus RNS- piesaistes domēnus un domēnus mijiedarbībai ar citiem proteīniem. Parasti SR proteīnu piesaiste pie ESE atvieglo eksona atpazīšanu, bet hnRNPs piesaiste to samazina.
 saimes proteīni un. heterogēnās ribonucleoproteīna daļiņas (hnRNPs). Cilvēka nRNP proteīni satur vienu vai vairākus RNS- piesaistes domēnus un domēnus mijiedarbībai ar citiem proteīniem. Parasti SR proteīnu piesaiste pie ESE atvieglo eksona atpazīšanu, bet hnRNPs piesaiste to samazina.")
68
pre-mRNA splaisings un slimības
Uzskata, ka visiem eksoniem ir šādi elementi. Mutācijas var ietekmēt regulatoros elementus un novest pie defektīva spalisinga. Ne tikai nesinonīmās mutācijas un stop kodoni, bet arī sinonīmās mutācijas var pārtraukt normālo splaisingu. Dažiem gēniem, līdz pat 50% punktveida mutācijas eksonos var ietekmēt splaisingu.

69
pre-mRNA splaisings un slimības
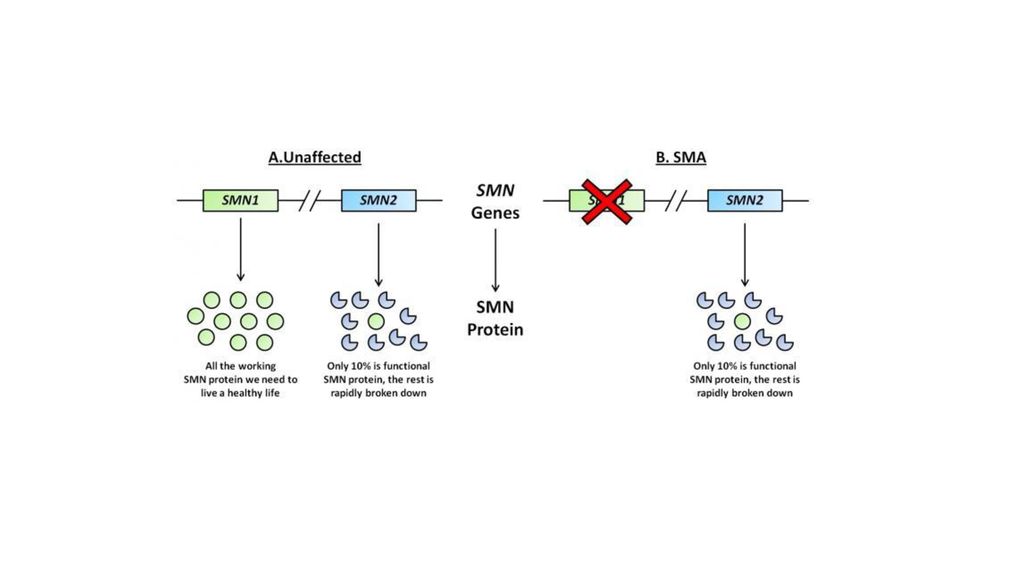
Mutācijas eksonu splaicinga vietās Mugurkaula muskulārā atrofija (spinal muscular atrophy (SMA) SMA ir samērā bieža (1 uz ), neiromuskulārā slimība. SMA ir raksturīga motoru neironu deģenerācija, kas izraisa progresējošu muskuļu vājumu. Slimību izraisa mutācija motoru neironu gēnā - SMN2. SMN gēns kodē proteīnu, kas spēlē būtisku lomu mazo nukleāro ribonukleoproteīnu (small nuclear ribonucleoproteins- snRNP) bioģenēzē.
70
pre-mRNA splicing and diseases
Spinal muscular atrophy (SMA) There are two SMN genes, SMN1 and SMN2, both of which encode the same open reading frame. The vast majority of SMA patients have deletions of the SMN1 gene . In the SMN2 gene they have a single nucleotide substitution, a C->T change at position 6 in exon 7. This mutation, though it does not change the amino acid coding, significantly alters the splicing pattern of the SMN2 pre-mRNA, causing frequent skipping of exon 7 that produces an inactive and unstable protein lacking the last 16 amino acids.
 There are two SMN genes, SMN1 and SMN2, both of which encode the same open reading frame. The vast majority of SMA patients have deletions of the SMN1 gene . In the SMN2 gene they have a single nucleotide substitution, a C->T change at position 6 in exon 7. This mutation, though it does not change the amino acid coding, significantly alters the splicing pattern of the SMN2 pre-mRNA, causing frequent skipping of exon 7 that produces an inactive and unstable protein lacking the last 16 amino acids.")
72
pre-mRNA splicing and diseases
Spinal muscular atrophy (SMA) Two models have been proposed to explain exon 7 skipping in SMN2: the substitution disrupts an ESE, which the splicing activator binds; it creates an ESS to which the splicing supressor hnRNP binds. Splicing is altered because of the balance of positively and negatively-acting splicing factors has been shifted.
 Two models have been proposed to explain exon 7 skipping in SMN2: the substitution disrupts an ESE, which the splicing activator binds; it creates an ESS to which the splicing supressor hnRNP binds. Splicing is altered because of the balance of positively and negatively-acting splicing factors. has been shifted.")
73
RNS splaisings Ir ļoti svarīgi lai introni tiktu izgriezti precīzi. Ja paliek pāri kāds introna nukleotīds vai tiek aizvākts kāds eksona nukleotīds, tas var novest pie nefunkcionāla proteīna sintēzes. Tas ir tāpēc, ka katru aminoskābes atlikumu kodē kodons, kas sastāv no 3 nukleotīdiem.

74
pre-mRNS splaisings Eksons: ” AMA NNA VKU RIE T”.
Neprecīza introna izgriešanas gadījumā veidojas lasīšanas rāmja nobīde, kā rezultātā būs pilnīgi jauna kodonu secība un veidosies nefunkcionāls proteīns. Lasīšanas rāmis attiecas uz kodonu secību, jo tie tiek lasīti no kāda specifiska sākumpunkta. Eksons: ”MANNAVKURIET”. ja introns tiek neprecīzi izgriezts, tad mēs vairs nevaram pareizi nolasīt eksonus: Eksons: ” AMA NNA VKU RIE T”.

75
Alternatīvais splaisings
Alternatīvais splaisings ir veids kā eksoni dažādās kombinācijās var savienoties viens ar otru, izveidojot dažādas nobriedušas mRNS (katrai būs iekodēta atšķirīga proteīna sekvence). Alternatīvais splaisings no ierobežota DNS daudzuma ļauj izveidot lielu proteinu dažādību. No 30 līdz 60% cilvēka gēnu ir pakļauti alternatīvam splaisingam. Daudzas slimības (15%) var būt saistītas ar nepareizi veikto alternatīvo splaisingu. Alternatīvais splaisings izskaidro kā – cilvēka genoma gēni var kodēt vairāk nekā dažādus mRNS molekulas.
. Alternatīvais splaisings no ierobežota DNS daudzuma ļauj izveidot lielu proteinu dažādību. No 30 līdz 60% cilvēka gēnu ir pakļauti alternatīvam splaisingam. Daudzas slimības (15%) var būt saistītas ar nepareizi veikto alternatīvo splaisingu. Alternatīvais splaisings izskaidro kā – cilvēka genoma gēni var kodēt vairāk nekā dažādus mRNS molekulas.")
76
Alternatīvais splaisings
Fibronektīns ir cilvēka gēna piemērs, kas iziet alternatīvo splaisingu. Vairāk nekā 20 dažādas fibronektīna izoformas tika atklātas.

77
RNS editēšana RNS editēšana ir jebkurš process, izņemot splaisingu, kura rezultātā mainās RNS transkripta sekvence un tā atšķiras no matricas DNS sekvences. pre-mRNS sekvences enzimātiska rediģēšana Bieži izplatīta vienšūņu mitohondrijos, kur vairāk kā 50% nukleotīdu sekvences var tikt izmainīta Daudz retāk sastopama augstākajos eikariotos Piemērs: H. sapiens apolipoproteīna gēna apoB pre-mRNS editēšana apob- apolipoprotein B in humans – part of large lipopr. complexes that transport lipids to serum. Only apob100 containing compl. transport cholesterol to body tissues via low density lipopr. complexes by binding to LDL receptors

78
RNS editēšanas mehānismi
1) Aizvietošanas editēšana Nukleotīdu ķīmiska izmainīšana - deaminēšana Piemēri: C deaminēšana par U vai A par I (inozīns, kuru ribosoma nolasa kā G) Reakciju veic deamināzes (piem. citozīna deamināze) 2) Ievietošanas/deletēšanas editēšana Uridīnu ievietošana/deletēšana
 Aizvietošanas editēšana. Nukleotīdu ķīmiska izmainīšana - deaminēšana. Piemēri: C deaminēšana par U vai A par I (inozīns, kuru ribosoma nolasa kā G) Reakciju veic deamināzes (piem. citozīna deamināze) 2) Ievietošanas/deletēšanas editēšana. Uridīnu ievietošana/deletēšana.")
79
RNS editēšana Cilvēka apolipoproteīna ApoB gēns kodē 14.1 kb mRNS transkriptu un 4536 aminoskābju produktu, ApoB 100. Zarnās specifisks enzīms- citozīna deamināze, APOBEC1, pārvērš citozīnu pozīcijā 6666 par uridīnu, izveidojot priekšlaicīgu stop kodonu. Saīsinātais mRNS kodē produktu, apoB48, kurš satur tikai pirmās 2152 aminoskābes no ApoB100 proteīna. Tikai pilna garuma ApoB100 zema blīvuma lipoproteīns var transportēt šūnā holesterolu. Cilvēka apoB pre-mRNS editēšana

80
Procesēta mRNS caur nukleārām
porām tiek transportēta uz citoplazmu, kur tālāk tiek translēta proteīnā.

81
Transkripcijas atšķirības prokariotu un eikariotu šūnās
Prokariotos transkripcija notiek citoplazmā. Turklāt transkripcijas un translācijas procesi notiek vienlaicīgi. Eikariotos transkripcija notiek šūnas kodolā, bet translācija citoplazmā. Līdz ar to abi procesi tiek atdalīti laikā un telpā.

82
Transkripcijas atšķirības prokariotu un eikariotu šūnās
Promoteri Prokariotu šūnā promoters sastāv no 2 īsām sekvencēm -10 un -35 pozīcijas no transkripcijas sākuma vietas . Sekvence -10 pozīcijā saucās Pribnow bokss, un parasti sastāv no 6 nukleotīdiem- TATAAT. Pribnow bokss ir absolūti nepieciešams transkripcijas iniciācijai. Otra sekvence -35 pozīcijā parasti sastāv no 6 nukleotīdiem - TTGACA. Šīs sekvences klātbūtne paIielina transkripcijas ātrumu. Eikariotu šūnā promoteri ir ļoti dažādi.

83
Transkripcijas atšķirības prokariotu un eikariotu šūnās
Enhānseri Eikariotu transkripciju pastiprina transkripciju regulējošie proteīni- enhānseri, kuri saistās ar DNS vietās, kas atrodas tālu no transkripcijas sākuma punkta. Prokariotu transkripcijas procesā nav enhānseru proteīnu.

84
Transkripcijas atšķirības prokariotu un eikariotu šūnās
RNS Pol RNS polimerāze būtībā veic vienu un to pašu reakciju visās šūnās. Prokariotu šūnās ir tikai viena RNS Pol, bet eikariotu šūnās tādas ir trīs- RNS Pol I, II un III. Prokariotu RNS Pol sastāv no 5 subvienībām (α,β,β’,ω). Eikariotu RNS Pol sastāv no 8-17 subvienībām.
. Eikariotu RNS Pol sastāv no 8-17 subvienībām.")
85
Transkripcijas atšķirības prokariotu un eikariotu šūnās
Eikariotu šūnā, histonu proteīna- DNS kompleksam ir jābūt pieejamam pirms transkripcijas procesa uzsākšanas. Prokariotu šūnās, DNS nav saistīta ar histonu proteīniem, līdz ar to transkripcija var notikt tieši.

86
Transkripcijas atšķirības prokariotu un eikariotu šūnās
Prokariotu šūnās transkripcijas laikā uzsintezēta mRNS netiek modificēta. Eikariotu šūnas modificē mRNS veicot RNS splaisingu, kepingu un poliadenilēšanu.

87
Translācija

Līdzīgas prezentācijas